Cataglyphis wird, wie alle Insekten, in Kopf, Thorax und Abdomen unterteilt.

Wissenschaftler sind uneinig über die Körpergliederung bei Ameisen, Abdomen wird manchmal in Petiolus und Gaster unterteilt.
Kopf im Vergleich zu anderen Ameisengattungen:
-relativ große Augen zur besseren Orientierung
-Fühler liegen tiefer, um den Blickwinkel nicht einzuschränken
-Mandibeln relativ klein, da keine Jäger, sondern Aasfresser
Komplexauge von Cataglyphis
Das Komplexauge von Cataglyphis ist, wie bei allen Ameisen, ein Appositionsauge
Appositionsaugen zeichnen sich durch durchgängige Pigmentzellen aus (Ommatidien ohne Zwischenräume)
Die Appositionsaugen bestehen bei Cataglyphis aus 500-1300 Ommatidien
Sichtbild ist ähnlich dem menschlichen, nur ist unseres viel feiner gerastert.
Lichtbrechungsapparat:
Cornealinse & Kristallkegel
Rhabdom:
besteht aus den Rhabdomeren von den 8 Sinneszellen, dient als Lichtwellenleiter
Pigmentzellen:
dienen zur Lichtabschirmung
Ocellen von Cataglyphis
Die Ocellen sind Medianocellen, sie liegen in der Kopfmitte
Ocellen sind bei Ameisen Becheraugen
Ocellen sind sehr simpel gebaut, sie können kein Bild erkennen
Die Aufgabe der Ocellen ist nicht zweifelsfrei geklärt!
Theorien:
Sie dienen zur Messung der Lichtstärke
Sie werden für eine Licht-Kompassorientierung verwendet.
Fühler von Cataglyphis
In Ameisenfühlern befindet sich der Tastsinn, der Geruchssinn und der Geschmackssinn.
Arten und Lebensraum
Bisher sind 65 Cataglyphis – Arten bekannt
Der bekannteste Cataglyphis – Forscher, R. Wehner, hat sich auf Cataglyphis fortis spezialisiert, die wir nun auch hier näher behandeln.
Cataglyphis fortis ist monogyn, sie haben nur eine Königin (identifizierbar am verdickten Mittelleib, wo sich früher die Flügel und die Flügelmuskulatur befand).
Cataglyphis fortis sind polymorph, es gibt keine „Soldaten“, sondern nur unterschiedlich große Arbeiter.
Eine Kolonie umfasst mehrere tausend, selten zehntausend Arbeiter.
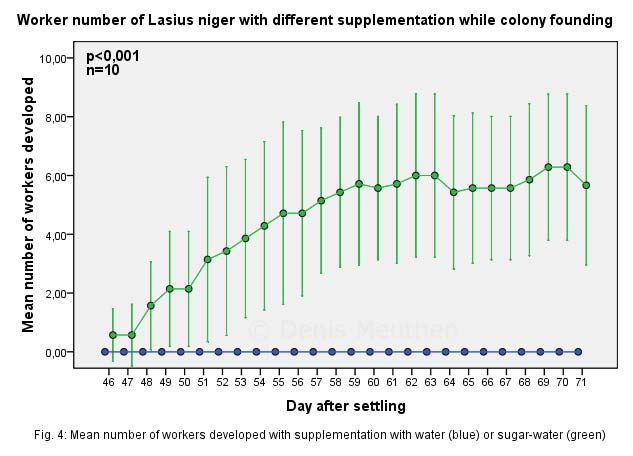
Die Entwicklung geht von Eiern über Larven zu Puppen bis zur fertigen Ameise (Holometabolie).
Ameisen wachsen nach dem Puppenstadium nicht weiter, ihre Größe wird schon im Larvenstadium durch die gegebene Futtermenge definiert.
Nach dem Ausschlüpfen aus der Puppenhülle halten sich die frischgeschlüpften, hellen Ameisen während dem Aushärteprozess bei der Königin auf, um ihren typischen „Nestgeruch“ anzunehmen.
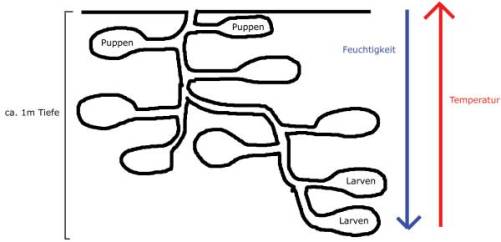
Cataglyphis fortis sind monodom, haben also nur ein Nest.
Der Bau ist hat nur einen Eingang und ist bis zu 1m tief.
Puppen befinden sich zum Ausbrüten in den wärmeren Kammern des Baus
Larven befinden sich in den feuchteren Kammern des Baus.
Die Königin bewegt sich durch den ganzen Bau, präferiert aber den feuchteren Teil.
Eier werden hinter der Königin hinterher getragen, also befinden sie sich meist auch im feuchteren Teil.
Cataglyphis fortis leben an einem Extremstandort: in der Sahara.
Vorteile:
Keine Prädatoren.
Keine konkurrierenden Arten, nur am Wüstenrand. Dort werden sie allerdings von anderen Arten verdrängt.
Nachteile:
Hohe Temperaturen, zu hoch für einen dauerhaften Aufenthalt außerhalb des Baus.
Wenige andere Lebewesen zum Beutefang vorhanden
Pheromone verflüchtigen sich durch hohe Temperatur schnell.
Wenige Landmarken.
Cataglyphis fortis suchen außerhalb ihres Baus hauptsächlich nach in der Hitze verendeten anderen Insekten.
Auch Samen und Früchte werden, falls vorhanden, gefressen.
Ihr Abdomen ist bei der Fortbewegung auf dem Sand meist nach oben geschwungen, damit es möglichst kühl bleibt und nicht überhitzt.
Je heißer es ist, desto kürzere Distanzen bewegen sie sich zur Nahrungssuche vom Nest fort. Die optimale Jagdzeit ist kurz nach Sonnenaufgang.
Nach Auffinden der Beute wird ihre Größe sowie ihre Distanz zum Nest abgewogen.
Große Beute wird nur nahe zum Nest von vielen Arbeitern in den Bau getragen
Kleine Beute auch von einzelnen Arbeitern bis zu 200 m entfernt vom Nest
Die Beute wird auf dem geographisch schnellstmöglichen Weg zurück ins Nest gebracht.
Wie schaffen sie diese Leistung?
Orientierung von Cataglyphis
1986 zeigten Wehner & Wehner in einem Experiment, dass Cataglyphis fortis nach einem Fund von Futter den geographisch schnellstmöglichen Weg zurück zum Nest nimmt.
Versetzt man die Ameise nach dem Futterfund in eine andere Gegend, läuft sie im gleichen Winkel die gleiche Distanz zu dem Ort, wo das Nest geographisch sein müsste. Die Versetzung wird also nicht registriert.
Schlussfolgerung: Für diese Leistung müssen die Ameisen 2 Mechanismen besitzen:
A) Eine Möglichkeit, sich zu orientieren, um den richtigen Winkel zurück zum Nest zu treffen.
B) Eine Möglichkeit, die Distanz zu messen.
Theoretische Überlegungen: Wegintegration
Schifffahrer verwenden seit Kenntnis der Mathematik die Vektor-Summen-Berechnung, um ihre Position auf See zu ermitteln. Dazu berechnen sie die Position aus der Summe der zurückgelegten Distanz und den Winkeln der zurückgelegten Strecken.
Ist ein solcher Mechanismus bei Cataglyphis vorhanden ? Können Ameisengehirne einen mittleren Laufwinkel anhand der Addition der Winkeländerungen relativ zum Sonnenstand über die Zeit berechnen?
Wehner zeigte 1983 den Gegenbeweis: Ameisen zeigen eine leichte Ungenauigkeit (Winkelabweichung (ε)) auf der Rückkehr zum Nest, je größer der Winkel α ist. Das wäre bei der Anwendung der Wegintegration aber nicht der Fall.
–> Mechanismus der Wegintegration bei Ameisen viel simpler gelöst
Zwischenthema: Was ist Polarisation / polarisiertes Licht?
Polarisiertes Licht = Lichtwelle, die in nur eine Richtung schwingt
Polarisiertes Licht entsteht durch Auftreffen von Sonnenstrahlen auf Partikel, die das Licht dann in die gleiche Richtung lenken. Teilpolarisiertes Licht entsteht also durch das Auftreffen von Licht auf die Atmosphäre (Streuung) oder auf Wasser (Reflexion).
Strahlung am stärksten in Richtung der Sonne –> Polarisationsrichtung dort am besten erkennbar
Der Winkel der Polarisation ändert sich mit unterschiedlicher Wellenlänge nur minimal. –> Lichtfarbe kaum Einfluss.
Polarisation als Orientierungsmöglichkeit
Vorteile:
Orientierung an polarisiertem Licht ist, als würde man einen Kompass verwenden. Die angegebene Richtung verändert sich nur bei Drehbewegungen, nicht bei linearen Bewegungen.
Die Änderungen des Lichtes über die Zeit sind vorhersagbar
Erste Ideen zur Orientierung am Licht kamen durch Santschi‘s Spiegelexperiment 1923 auf, in dem er zum Nest zurückkehrenden Ameisen mit Spiegeln einen falschen Sonnenstand vortäuschte und diese sich dann zum neuen Sonnenstand hin ausrichteten. Orientierung an einem Reiz ohne Aufgabe der Richtungsänderung = Menotaxis
Erst ca. 60 Jahre später, 1982 zeigte Wehner mit Hilfe einer rollenden Apparatur in ihrem Lebensraum eindeutig, dass Cataglyphis in der Lage ist, sich anhand der Polarisationsrichtung des Lichtes zu orientieren.
Weitere Beweise für die Orientierung von Cataglyphis anhand der Polarisation
Schon 1976 entdeckte Wehner et al., dass sich in der dorsalen Augenregion von Cataglyphis fortis wie bei Bienen (1975) spezialisierte Photorezeptoren befinden, die zur Erkennung von polarisiertem Licht benötigt werden.
2006 entdeckte Ronacher et al., dass vor UV-Licht geschützte Bereiche (grauer Bereich in der Röhre) die Ameisen daran hindern, ihre Orientierung entsprechend zum Weg zu ändern –> polarisiertes Licht zur Orientierung stammt hauptsächlich aus dem UV-Bereich
Aufbau der Ommatidien in der dorsalen Augenregion bei Cataglyphis
1993 untersuchten Labhart & Meyer elektronenmikroskopisch die „dorsal rim area“ von Cataglyphis bicolor
Die Rhabdome in der DRA haben charakteristische Formen. Auch besitzen sie nur 2 Gegenden, in denen die Mikrovili senkrecht laufen. Die Mikrovili sind in einem 90°-Winkel zueinander ausgerichtet.
Rhabdomere der UV-Rezeptoren bilden den größten Teil der Zellen in der DRA = Polarisation wird vor allem im UV-Bereich wahrgenommen
Aber: Es wurde nachgewiesen, dass Cataglyphis sich auch mit einer abgedeckten DRA grob an der Polarisation orientieren können!
–> Normale Ommatidien erkennen auch polarisiertes Licht im UV-Bereich
Landmarken
Nach der Versetzung parallel zum Nest läuft Cataglyphis Orienteriungsschleifen um die eigentliche Nestposition. Findet sie aber ein bekanntes Bild einer Landschaft, so findet sie den schnellstmöglichen Weg zur tatsächlichen Nestposition.
1997 wies Akesson et al. nach, dass Cataglyphis sich zur Orientierung, wie zuvor schon bei anderen Insekten gezeigt wurde, auch an Landmarken mit Hilfe von „Schnappschusskarten“ orientieren können.
„Schnappschusskarten“ = vereinfachtes, zweidimensionales Bild von Landmarken
Beim Betrachten einer neuen Gegend: Vergleich mit eingespeicherten Schnappschusskarten zur Orientierung, größte Übereinstimmung wird gewählt.
2002 zeigte Akesson et al., dass Cataglyphis in der Lage ist, „Schnappschusskarten“ zumindest temporär unabhängig von der Polarisation als Orientierung zu benutzen. Andere Wüstenameisenarten benutzen ausschließlich Landmarken zur Orientierung.
Im gleichen Jahr zeigte Collet et al., dass die Erkennung von Landmarken den Ameisen nicht ihre Position verrät, sondern die Erkennung der Landmarke mit der darauf folgenden Aktion gekoppelt ist.
Gerüche
1999 fanden Wolf & Wehner, dass Cataglyphis auch Gerüche zur Orientierung verwenden. Werden von einem Kadaver Gerüche emittiert, so folgt Cataglyphis den Geruchswolken und nicht dem direkten Weg zur Beute.
2005 entdeckten die beiden Forscher, dass die Orientierung nach den Geruchswolken (und damit bei Südwind eine anfängliche Orientierung weg von der Beute) vorteilhaft ist, da ein direkter Weg vor allem bei großen Distanzen einen hohen Fehler und damit eine hohe Ungenauigkeit aufweisen würde.
Distanzmessung
Cataglyphis fortis ist in der Lage, die exakte Position nicht nur durch die Richtung zum Nest, sondern auch durch die Entfernung zu ermitteln. Menschen ermitteln die gelaufene Distanz anhand der Zeit. Wie aber löst das Cataglyphis?
2000 stellten Ronacher et al. fest, dass Cataglyphis fortis die Distanzmessung unabhängig von der visuellen Orientierung vollziehen. Selbst das Erkennen von Landmarken brachte die Ameisen nicht dazu, die zurückgelegte Distanz zu verändern.
2003 fanden Sommer & Wehner heraus, dass Cataglyphis fortis dazu neigen, die zurückgelegte Distanz vor allem bei langen Wegen leicht zu unterschätzen.
Lange Zeit blieb diese Frage offen, bis 2006 Wittlinger et. al. den Ameisen nach Futterfund die Beine verkürzten oder mit Schweineborsten verlängerten und entdeckten, dass sie die Distanz zurück zum Nest unter- bzw. überschätzen.
–>Cataglyphis fortis besitzen einen neuronalen Odometer (Schrittzähler)
Kehrt Cataglyphis fortis mit manipulierten Beinen zurück zum Nest, so ist sie bei der nächsten Beutesuche wieder in der Lage, die Distanz korrekt einzuschätzen –> „Schrittzähler“ wird zurückgesetzt. Ohne zu wissen, dass es sich beim Distanzmesser um ein Odometer handelt, stellten Knaden & Wehner diesen Fakt schon 2005 fest.
Die dritte Dimension wird beim Schrittzähler mit einbezogen, die aufgenommene Distanz berechnet also die Summe der Auf- und Abstiege mit ein. Das konnte 2002 durch Wohlgemuth & Ronacher & Wehner dadurch gezeigt werden, dass die Ameisen selbst bei Auf- und Abstiegen in der Lage waren, die Distanz zum Nest zurück korrekt zu ermitteln. 2005 bestätigten Grah & Wehner & Ronacher diese Leistungen von Cataglyphis fortis.
Sahabot – ein Roboter mit Cataglyphis-Leistungen
Mit Hilfe von R. Pfeifer, einem Informatiker, stellte R. Wehner 1996 einen Roboter namens Sahabot her, der in der Lage war, sich mit den gleichen Orientierungsmöglichkeiten von Cataglyphis fortis zu orientieren: am polarisierten Licht und später auch an Landmarken mittels „Schnappschusskarte“.
Die erste Version erlaubte nur eine Orientierung nach Polarisation und benötigte noch einen externen Ständer. Die 1999 entwickelte zweite Version integrierte die Polarisationserkennung in den Roboter selber und fügte eine Möglichkeit hinzu, sich anhand einer Schnappschusskarte zu orientieren.
Weitwinkel-Sensoren imitieren die im Insektengehirn befindlichen polarisationsempfindlichen Grossfeldneuronen, eine Kamera fotografiert Landmarken. Ein interner Prozessor gleicht die Landmarken mit bereits gespeicherten Bildern ab.
Der Roboter erlaubte, die Theorien über die mögliche Orientierung von Cataglyphis fortis anhand der Polarisation und anhand einer Schnappschusskarte zu beweisen. Auch die Funktion des der erst 2006 entdeckten Schrittzählers könnte leicht anhand Sahabot bewiesen werden.