- 15.04.2026 Trends in Ecology and Evolution on the interplay between individualisation and epigenetics well received in national and international press
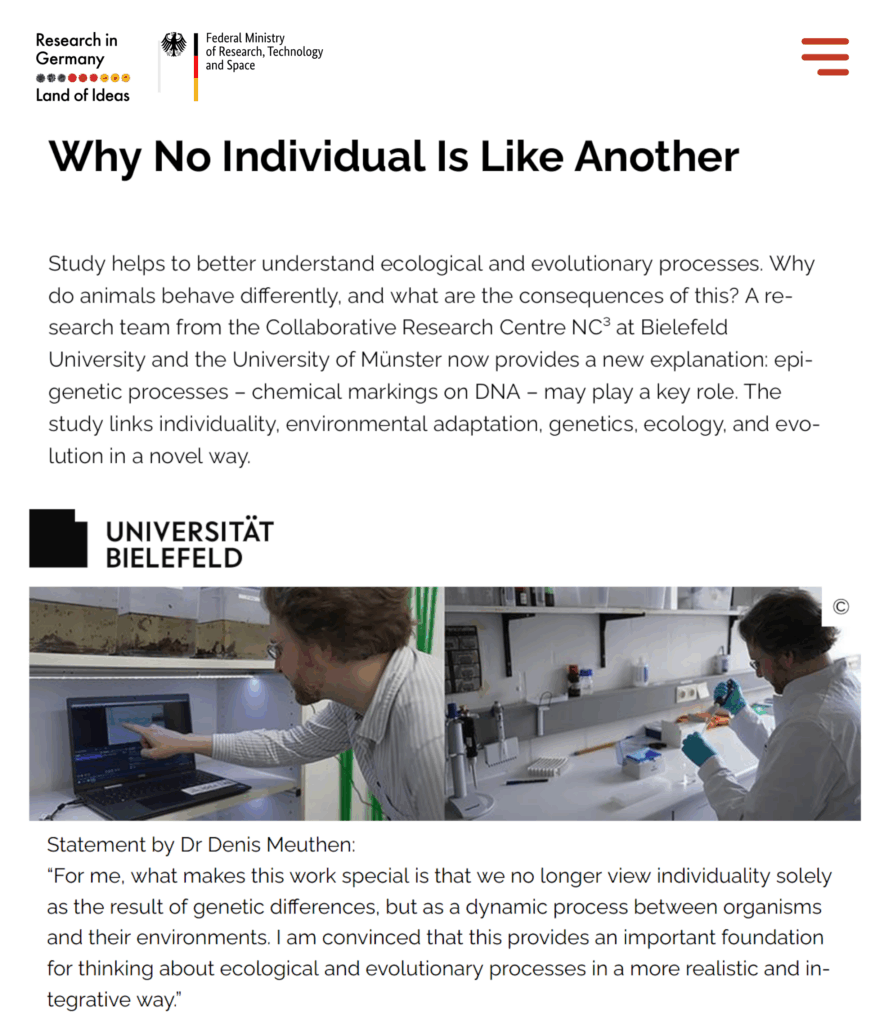
- 24.03.2026 Paper on the interplay between individualisation and epigenetics published in Trends in Ecology and Evolution
- 12.08.2025 Paper on risk allocation in Physella acuta published in Behavioral Ecology!
- 05.07.2025 Paper on the effects of long-term gastropod alarm cue frozen storage published in Animal Behaviour!
- 01.07.2025 Paper on the consequences of citalopram on gastropod antipredator plasticity published in Ecotoxicology and Environmental Safety!
Paper on the interplay between individualisation and epigenetics published in Trends in Ecology and Evolution
Paper on risk allocation in Physella acuta published in Behavioral Ecology!
Paper on the effects of long-term gastropod alarm cue frozen storage published in Animal Behaviour!
Paper on the consequences of citalopram on gastropod antipredator plasticity published in Ecotoxicology and Environmental Safety!
Career column published in Trends in Ecology and Evolution
Preprint on the interplay of epigenetics and individualization now available on EcoEvoRxiv!
Paper on the integration of transgenerational morphological plasticity published in Functional Ecology!
“Outstanding new principal investigator” awarded!

Today, I received the award of an “outstanding new principal investigator (PI)” by a worldwide community of over 2500 new principal investigators (i.e., assistant professors). I’m proud of being the first person from Germany and the second person in Europe to have received this award to date. I was elected due to being an example for diversity in research (see my Nature and Science articles), for having a very good publication output (11 publications) since my start as a Freigeist Fellow in 2022, and for my helpful and valuable efforts to support other new PI’s to tackle common challenges. In the interview that accompanies this award, I share my research focus, my challenges, and my strategies in leading my own workgroup.
Photo: © S.Jonek/Bielefeld University
Working Life column published in Science

Today, my Working Life piece on the strategies that I use to lead an academic workgroup as a lip-reading scientist was published in Science. The strategies that I share in this article should be helpful for any principal investigator, disabled or not.
Photo: © SereDim/ndla.no